Нейроны. Их классификация, физиологические свойства, связь с нейроглией. Распространение возбуждения по нервным волокнам. Характеристика их возбудимости и лабильности. Нейронная теория
А)Возбудимость- способность живых клеток воспринимать изменения внешней среды и отвечать на эти изменения (раздражения) реакцией возбуждения.
Б)Рефрактерность- невозбудимость во время возбуждения
В) Лабильность- максимальный ритм раздражений, который живая ткань может воспроизвести за еденицу времени.
Г)Сократимость- Способность развивать силу и напряжение, изменяя конфигурацию, при переходе в состояние возбуждения.
Д) проводимость- Способность клеток органов и тканей проводить волну возбуждения.
Морфофункциональные особенности биологических мембран
Все мембраны по своей организации и составу обнаруживают ряд общих свойств. Они:
состоят из липидов, белков и углеводов;
являются плоскими замкнутыми структурами;
имеют внутреннюю и внешнюю поверхности (асимметричны);
избирательно проницаемы.
Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойствомамфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду.
Мембранный потенциал покоя и его происхождение
Мембранным потенциалом покоя (МПП) или потенциалом покоя (ПП) называют разность потенциалов покоящейся клетки между внутренней и наружной сторонами мембраны.
Основные положения мембранной теории происхождения МПП сводятся к следующему. В состоянии покоя клеточная мембрана хорошо проницаема для ионов К + (в ряде клеток и для СГ), менее проницаема для Na + и практически непроницаема для внутриклеточных белков и других органических ионов. Ионы К + диффундируют из клетки по концентрационному градиенту, а непроникающие анионы остаются в цитоплазме, обеспечивая появление разности потенциалов через мембрану.
Возникающая разность потенциалов препятствует выходу К + из клетки и при некотором ее значении наступает равновесие между выходом К + по концентрационному градиенту и входом этих катионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется равновесным потенциалом.
Мембранный потенциал действия и его фазы. Соотношение фаз возбудимости с фазами ПД
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания.
Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Быстрое и большое количество поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит.
Потенциал действия (ПД)-быстрое колебание МП-самораспространяющийс процесс,связанный с изменениями ионной проводимости мембраны,вызванным функционированием ионных каналов. ПД распространяется без затухания,т е практически без уменьшения амплитудыМежду моментом нанесения раздражения и первым проявлением ПД имеется задержка-латентный период(ЛП). ЛП соответствует времени,когда ПД движется по мембране нервной клеткиот места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны-локальный ответ.При достижении критического уровня деполяризации,сост в средн-55 мВ,начинается фаза деполяризации.В эту фазу уровень МП падает до 0 и даже преобретает положительное значение (овершут),а затем возвращается к исходному уровню фаза реполяризации.Фазы деполяризации,овершута и реполяризации образуют спайк (пик)ПД,длительность которого(1-2 милисек).После спайка наблюдается замедление скорости спада потенциала-фаза следовой деполяризации.После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации.Они могут длиться десятки,сотни миллисек.В основе изменений МП,происходящий в течении ПД,лежат ионные механизмы.Локальный потенциал.раздражение клетки приводит к открытию части Na+каналов и появлению локального (нераспространяющегося потенциала)Фаза деполяризации.При достижении критического уровня деполяризации мембраны начинается лавинообразный процесс открытия большлго количества Na+-каналов.В фазу деполяризац происходит массивный вход в клетку ионов Na+по концетрационному и электрохимическому градиенту.Овершут.Деполяризация мембраны приводит к реверсии МП(МП станов положит).В эту фазу Na+-ток начинает стремительно спадать,что связано с инактивацией потенциало-зависимых Na+-каналов(время открытого состояния-доли сек) и исчезновением электрохимического градиента Na+.Фаза реполяризации.Инактивация Na+-каналов открытие потенциалозависимых К+-каналов способствуют развитию реполяризации.Выход К+ наружу способствует полноценному завершению фазы реполяризац.Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов.Восстановление исходного уровня Мп приводит Na- и К-каналы в состояние покоя.В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки.Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течении которого даже сверхпороговое раздражение не способно вызвать пД.Это связано с инактивацией большинства Na-кан.Относительно рефрактерный период.В конце фазы реполяризации,а так же во время следовой гипераполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители.Это связано со значительным реполяризующим действием выходящих Ка токовЛабильность-максимально возможная частота генерации ПД для данного типа возбудимой клетки.Лабильность большинства нейронов составляет приблиз-но 400 ПД/сСоотношение фаз возбудимости с фазами ПДУровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности.В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности.Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости. Следует отметить, что явление аккомодации также связано с изменением проводимости ионных каналов. Если деполяризующий ток нарастает медленно, то это приводит к частичной инактивации натриевых, и активации калиевых каналов. Поэтому развития ПД не происходит.Возбуждение может быть 2-х видов: местное (локальный ответ);распространяющееся (импульсное). Местное возбуждение- наиболее древний вид (низшие формы организмов и низковозбудимые ткани - например, соединительная ткань). Местное возбуждение возникает и в высокоорганизованных тканях под действием подпорогового раздражителя или как компонент потенциала действия. При местном возбуждении нет видимой ответной реакции. Особенности местного возбуждения: нет латентного (скрытого) периода - возникает сразу же при действии раздражителя; нет порога раздражения; местное возбуждение градуально - изменение заряда клеточной мембраны пропорционально силе подпорогового раздражителя; нет рефрактерного периода, наоборот характерно небольшое повышение возбудимости; распространяется с декрементом (затуханием). Импульсное (распространяющееся) возбуждение - присуще высокоорганизменным тканям, возникает под действием порогового и сверхпорогового раздражителей.Особенности импульсного возбуждения: имеет латентный период - между моментом нанесения раздражения и видимой ответной реакцией проходит некоторое время; имеет порог раздражения; не градуально - изменение заряда клеточной мембраны не зависит от силы раздражителя; наличие рефрактерного периода; импульсное возбуждение не затухает. Вывод:в организме животного и человека наблюдается местное и импульсное возбуждение. Возникновение того или иного вида возбуждения зависит от степени развития ткани и силы раздражителя.
Закон силы раздражения
Чем больше сила раздражения, тем больше, до известных пределов, величина ответных реакций.
Закон силы времени
Чем больше сила раздражителя, тем
меньше по времени может действовать
данный раздражитель для получения
ответной реакции.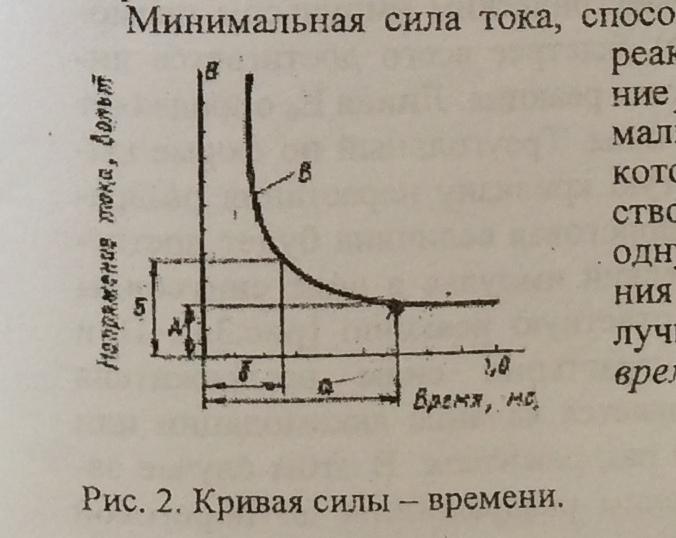
Закон градиента раздражения
Чем быстрее нарастает сила раздражителя
до пороговой величины, тем быстрее
появляется ответная реакция.
Нейрон, его физиологические свойства, классификация, особенности возникновения и распространения возбуждения
Нейрон (от др.-греч. νεῦρον - волокно, нерв) - это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высокоспециализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов. Место контакта двух нейронов называется синапсом.
Нейрон состоит из тела диаметром от 3 до 100 мкм, содержащего ядро и другие органеллы, и отростков. Выделяют два вида отростков. (Аксон - длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты - как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов, но только один аксон. Один нейрон может иметь связи с до 20 тысяч других нейронов.
Физиологические свойства
Нейроны восприимчивы к раздражению, то есть способны воспринимать раздражитель и отвечать на него генерацией потенциала действия (ПД). Обычно раздражителем для нейрона служит нейромедиатор, выделяемый другими нейронами в синаптические щели. Одни синапсы вызывают деполяризацию нейрона, другие - гиперполяризацию; первые являются возбуждающими, вторые - тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Классификация
По положению в рефлекторной дуге различают афферентные нейроны (сенсорные нейроны), эфферентные нейроны (двигательные нейроны) и вставочные нейроны.
На основании числа и расположения отростков нейроны делятся на униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные нейроны.
Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей. Этими особенностями являются следующие.
1. Иррадиация (дивергенция) возбуждения в ЦНС. Она объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся (рис. 4.4, а).
Иррадиацию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное энергичные движения всех конечностей и даже туловища. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
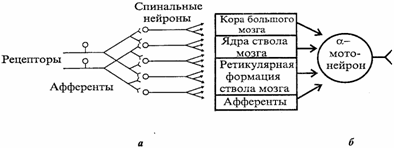
Рис. 4.4. Дивергенция афферентных дорсальных корешков на спинальные нейроны, аксоны которых, в свою очередь, ветвятся, образуя многочисленные коллатерали (в), и конвергенция эфферентных путей от различных отделов ЦНС на α-мотонейрон спинного мозга (6)
1. Конвергенция возбуждения (принцип общего конечного пути)схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу(принцип шеррингтоновской воронки).Конвергенция возбуждения объясняется наличием многих аксонных коллатералей,вставочных нейронов, а также тем, что афферентных путей в несколько раз больше,чем эфферентных нейронов. На одном нейроне ЦНС может располагаться до 10 000синапсов. Явление конвергенции возбуждения в ЦНС имеет широкое распространение.Примером может служить конвергенция возбуждения на спинальном мотонейроне. Так,к одному и тому же спинальному мотонейрону подходят первичные афферентные волокна (рис. 4.4, б), а также различные нисходящие пути многих вышележащих центров ствола мозга и других отделов ЦНС. Явление конвергенции весьма важно:оно обеспечивает, например, участие одного мотонейрона в нескольких различных реакциях. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг. На рис. 4.4, я показаны два афферентных волокна, каждое из которых отдает коллатерали к 4 нейронам таким образом, что 3нейрона из общего их числа, равного 5, образуют связи с обоими афферентными волокнами. На каждом из этих 3 нейронов конвергируют два афферентных волокна.
На один мотонейрон может конвергировать множество коллатералей аксонов, до 10 000-20 000, поэтому генерация ПД в каждый момент зависит от общей суммы возбуждающих и тормозящих синаптических влияний. ПД возникают лишь в том случае, если преобладают возбуждающие влияния.Конвергенция может облегчать процесс возникновения возбуждения на общих нейронах в результате пространственной суммации подпороговых ВПСП либо блокировать его вследствие преобладания тормозных влияний (см. раздел 4.8).
3. Циркуляция возбуждения по замкнутым нейронным цепям. Она может продолжаться минуты и даже часы (рис. 4.5).
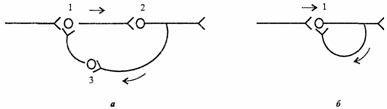
Рис. 4.5. Циркуляция возбуждения в замкнутых нейронных цепях по Лоренто де-Но (а) и по И.С.Беритову (б). 1,2,3возбуждающие нейроны
Циркуляция возбуждения одна из причин явления последействия, которое будет рассмотрено далее (см. раздел 4.7). Считают, что циркуляция возбуждения в замкнутых нейронных цепях наиболее вероятный механизм феномена кратковременной памяти (см. раздел 6.6). Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом.
4. Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах. Распространение возбуждения от аксона одного нейрона к телу или дендритам другого нейрона, но не обратно объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении(см. раздел 4.3.3).
5. Замедленное распространение возбуждения в ЦНС по сравнению с его распространением по нервному волокну объясняется наличием на путях распространения возбуждения множества химических синапсов. Время проведения возбуждения через синапс затрачивается на выделение медиатора в синаптическую щель, распространение его до постсинаптической мембраны,возникновение ВПСП и, наконец, ПД. Суммарная задержка передачи возбуждения в синапсе достигает примерно 2 мс. Чем больше синапсов в нейрональной цепочке,тем меньше общая скорость распространения по ней возбуждения. По латентному времени рефлекса, точнее по центральному времени рефлекса, можно ориентировочно рассчитать число нейронов той или иной рефлекторной дуги.
6. Распространение возбуждения в ЦНС легко блокируется определенными фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов.
Рассмотренные особенности распространения возбуждения дают возможность подойти к пониманию свойств нервных центров.
Классификация физиологические свойства и функции нервных волокон
Нервные волокна представляют собой отростки нейронов, с помощью которых осуществляется связь между нейронами, а также нейронов с исполнительными клетками. В состав нервного волокна входят осевой цилиндр (нервный отросток) и глиальная оболочка. По взаимоотношению осевых цилиндров с глиальными клетками выделяют два типа нервных волокон: безмиелиновые и миелиновые. Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты). При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полностью окружает каждый осевой цилиндр и смыкается над ним, образуя сдвоенную мембрану (мезаксон
Наиболее распространена классификация по Дж.Эрлангеру и Х.Гассеру (1937), в которой волокна разделяют на три типа: А, В и С (табл.3). Волокна типа А и В являются миелиновыми, типа С - безмиелиновыми. Волокна А делят на 4 подгруппы: α, β, γ, δ. В периферической нервной системе к волокнам Аα относятся афферентные волокна от механорецепторов кожи, мышечных и сухожильных рецепторов, а также эфферентные волокна к скелетным мышцам. К Аβ принадлежат афферентные волокна от кожных рецепторов прикосновения и давления, от части мышечных и висцеральных рецепторов. Аγ представляют собой эфферентные волокна, через которые регулируется активность мышечных рецепторов. К Аδ относят афферентные волокна от части тактильных, температурных и болевых, а также суставных рецепторов. К волокнам типа В принадлежат преганглионарные волокна вегетативной нервной системы. К волокнам типа С относят постганглионарные волокна вегетативной нервной системы, афферентные волокна от некоторых болевых (вторичная боль), тепловых и висцеральных рецепторов.
Таблица 3. Типы волокон в нервах млекопитающих (по Эрлангеру-Гассеру)
|
Тип волокон |
Диаметр волокна, мкм |
Скорость проведения возбуждения, м/с |
Длительность абсолютного рефракторного периода, мс |
Из данных, представленных в табл.3, видно, что средний диаметр каждого типа волокна снижается от типа А до С (каждый примерно в 2 раза по отношению к предыдущему). Соответственно этому снижается и скорость проведения возбуждения. Низкая скорость проведения нервного импульса в волокнах типа С связана с особенностями проведения возбуждения в безмиелиновых волокнах. Лабильность также уменьшается от волокон Аα до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость тоже уменьшается от волокон Аα (наибольшая возбудимость) к волокнам С (наименьшая возбудимость). Например, пороговая сила электрического тока у волокон С в 30-50 раз больше, чем у волокон Аα. Исследование факторов, блокирующих нервную проводимость, показало, что к давлению наиболее чувствительны волокна А, к кислородному голоданию (гипоксии) - волокна В, к местным анестетикам - волокна С.
Нервные волокна имеют две основные функции - проведение возбуждения и транспорт веществ, обеспечивающих трофическую функцию
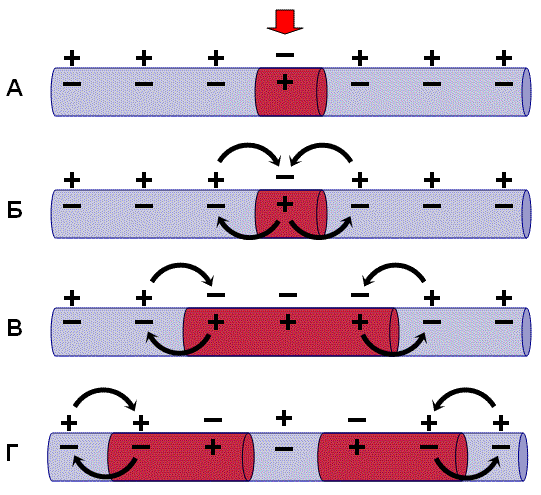
Законы проведения возбуждения в нервных стволах
^ Закон физиологической и анатомической непрерывности - возбуждение может распространяться по нерву только при сохранении его морфологической и функциональной целостности. Травматическое повреждение нерва нарушают или полностью прекращают проведение возбуждения.
^ Закон изолированного проведения - возбуждение, распространяющееся в одной группе волокон (например, A«), не передается на волокна другой группы (например, В) того же ствола. Вследствие этого информация, передаваемая по разным типам волокон, носит строго направленный специфический характер.
^ Закон двустороннего проведения - возбуждение, возникающее в каком-либо участке нерва, распространяется в обе стороны от очага возникновения.
функции нервной системы рефлекторный принцип ее деятельности. Анализ структуры рефлекторной дуги.
Нервная система человека делится на центральную (головной и спинной мозг) и периферическую. Центральная нервная система обеспечивает индивидуальное приспособление, поведение организма в конкретных условиях среды обитания, регулирует деятельность каждого органа, обеспечивает интеграцию и объединение органов в единую систему, согласует интенсивность функционирования систем организма, обеспечивает реагирование организма как единого целого на раздражители из внешней и внутренней среды организма. Общей функцией ЦНС является ее трофическое влияние на клетки. В органах, лишенных связей с ЦНС, развиваются дистрофические, воспалительные и атрофические процессы, приводящие к снижению и прекращению функциональной активности органов.
Рефлекс – закономерная реакция организма на изменение внешней и внутренней среды, осуществляемая при участии нервной системы в ответ на раздражение рецепторов. В процессе рефлекторной реакции воспроизводится, изменяется интенсивность или прекращается деятельность тканей, органов или организма в целом. При помощи рефлекса устанавливается адекватное соотношение активности органов в пределах системы, систем в пределах организма, организма в его взаимоотношениях с окружающей средой. Рефлекторный ответ осуществляется за минимальное время и с максимальной безошибочностью.
Морфологическим субстратом рефлекса является рефлекторная дуга. Ее звенья:
1. Афферентное (рецепторы и афферентный нейрон).
2. Центральное (вставочные нейроны и синапсы).
3. Эфферентное (эффекторный нейрон и эффектор).
Простейшая (моносинаптическая) рефлекторная дуга имеет два нейрона: афферентный и эфферентный и один синапс. Рефлекторные дуги большинства рефлексов полисинаптические.
Область тела, раздражение которой вызывает определенный рефлекс, называется рецептивным полем рефлекса (рефлексогенной зоной). Нервный центр – совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции той или иной функции. Время от начала раздражения рецептора до появления ответной реакции называется латентным периодом рефлекса. Более медленное проведение возбуждения по рефлекторной дуге, чем по нерву, связано с явлением синаптической задержки, необходимой для:
1) выделения медиатора нервным окончанием в ответ на импульс;
2) диффузии медиатора через синаптическую щель к постсинаптической мембране;
3) возникновения возбуждающего постсинаптического потенциала. Вместе с формированием пикового потенциала на мембране нейрона это время составляет 1,5 – 2,0 мс. Время, необходимое для проведения возбуждения по центральной части рефлекторной дуги (с аффекторных нейронов на эффекторные), зависит от количества вставочных нейронов и называется центральным временем рефлекса .
Классификация рефлексов
По биологическому значению : пищевые, половые, оборонительные, локомоторные, позно-тонические, ориентировочные.
В зависимости от расположения рецепторов : экстрарецептивные, интеррецептивные и проприорецептивные.
В зависимости от того, какие отделы мозга необходимы для осуществления рефлекса: спинальные, бульбарные, мезенцефальные, кортикальные.
В зависимости от отдела нервной системы, который реализует ответ: соматические или вегетативные.
По характеру ответной реакции : моторные, секреторные, сосудодвигательные. Моторные рефлексы по длительности ответной реакции разделяются на фазические и тонические.
По приспособительному значению рефлексы делятся на безусловные и условные.
Синапсы ЦНС, строение, свойства, классификация .
Синапс - специализированный контакт между нервными клетками или нервными клетками и другими возбудимыми образованиями, обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов осуществляется взаимодействие разнородных по функциям тканей организма, например нервной и мышечной, нервной и секреторной.^ Структура синапса .Пресинаптическое окончание аксона нейрона при подходе к иннервируемой клетке теряет миелиновую оболочку, что несколько снижает скорость распространения волны возбуждения. Небольшое утолщение на конце волокна, называемоесиноптической бляшкой, содержит синаптические пузырьки размером 20-60 нм с медиатором - веществом, способствующим передаче возбуждения в синапсе.^ Синаптическая щель - пространство между пресинаптическим окончанием и участком мембраны эффекторной клетки является непосредственным продолжением межклеточного пространства.^ Постсинаптическая мембрана - участок эффекторной клетки, контактирующий с пресинаптической мембраной через синаптическую щель.Классификация синапсов .В соответствии с морфологическим принципом синапсы подразделяют на: аксо-аксональные (между двумя аксонами); аксодендритические (между аксоном одного нейрона и дендритом другого); аксосоматические (между аксоном одного нейрона и телом другого); дендродендритические (между дендритами двух или нескольких нейронов); нервно-мышечные (между аксоном мотонейрона и исчерченным мышечным волокном); аксоэпителиальные (между секреторным нервным волокном и грану-лоцитом); межнейронные (общее название синапсов между какими-либо элементами двух нейронов).^ Все синапсы делят на центральные (в головном и спинном мозге) и периферические (нервно-мышечные, аксоэпителиальные и синапсы вегетативных ганглиев).В соответствии с нейрохимическим принципом синапсы классифицируют по виду химического вещества - медиатора, с помощью которого происходит возбуждение и торможение эффекторной клетки.^ По способу передачи возбуждения синапсы подразделяют на три группы. Первую составляют синапсы с химической природой передачи посредством медиаторов (например, нервно-мышечные); вторую - синапсы с передачей электрического сигнала непосредственно с пре- на постсинаптическую мембрану. Третья группа представлена «смешанными» синапсами, сочетающими элементы как химической, так и электрической передачи.По конечному физиологическому эффекту , а также по изменению потенциала постсинаптической мембраны, различают возбуждающие и тормозные синапсы.^ Механизм проведения возбуждения в синапсах. Передача возбуждения в химическом синапсе - сложный физиологический процесс, протекающий в несколько стадий. Он включает синтез и секрецию медиатора; взаимодействие медиатора с рецепторами постсинаптической мембраны; инактивирование медиатора. В целом синапс осуществляет последовательную трансформацию электрического сигнала, поступающего по нервному волокну, в энергию химических превращений на уровне синаптической щели и постсинаптической мембраны, которая затем снова трансформируется в энергию распространяющегося возбуждения в эффекторной клетке.Свойства синапсов .
Пластичность синапса .
Одностороннее проведение возбуждения.
Низкая лабильность и высокая утомляемость синапса обусловлены временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности.
Высокая избирательная чувствительность синапса к химическим веществам обусловлена специфичностью хеморецепторов постсинаптической мембраны.
Способность синапса трансформировать возбуждение связана с его низкой функциональной лабильностью и спецификой протекающих в нем химических процессов.
^ Синаптическая задержка , т.е. время между приходом импульса в преси-наптическое окончание и началом ответа, составляет 1-3 мс.Суммация возбуждений определяется переходом местного возбуждения в распространяющееся в результате временного взаимодействия серии возбуждающих постсинаптических потенциалов.
^ Трофическая функция синапсов
Медиаторные механизмы передачи возбуждения в ЦНС
Медиаторами, или нейротрансмиттерами, нейронов ЦНС являются различные биологически активные вещества. В зависимости от химической природы их можно разделить на 4 группы: 1) амины (ацетилхолин, норадреналин, дофамин, серотонин), 2) аминокислоты (глицин, глутаминовая, аспарагиновая, гамма-аминомасляная - ГАМК), 3) пуриновые и нуклеотиды (АТФ); 4) нейропептиды (вещество Р, вазопрессин, опоидни пептиды и др.). Раньше считали, что во всех окончаниях одного нейрона "выделяется один медиатор (по принципу Дейла). За последние годы выяснили, что во многих нейронах может содержаться 2 медиаторы или больше. По действию медиаторы можно разделить на ионотропных и метаболотропни. Ионотропных медиаторы после взаимодействия с циторецепторамы постсинаптической мембраны изменяют проницаемость ионных каналов. Метаболотропни медиаторы постсинаптическую действие проявляют путем активации специфических ферментов мембраны. Вследствие этого в мембране или (чаще) в цитоплазме клетки активируются так называемые вторичные посредники (вторичные мессенджеры), которые в свою очередь запускают каскады внутриклеточных процессов, тем самым влияя на функции клеток. К основным мессенджеров систем внутриклеточной сигнализации относят аденилатциклазной и полифосфоинозитидну. В основе первой лежит аденилатциклазной механизм. Центральным звеном второй системы является кальциймобилизуючий каскад полифосфоинозитидив, который контролируется фосфолипазой С. Физиологический эффект этих систем осуществляется путем активации специфических ферментов - протеинфосфокиназ, конечным итогом чего является широкий спектр воздействия на белковые субстраты, которые могут подвергаться фосфорилированию. Вследствие этого изменяется проницаемость мембран для ионов, синтезируются и выделяются медиаторы, регулируется синтез белков, осуществляется энергетический обмен и т.д.. Метаболотропним эффектом обладают большинство нейропептидов. Метаболические изменения, происходящие в клетке или на ее мембране под действием метаболотропних медиаторов, длительные, чем при действии ионотропных медиаторов. Они могут затрагивать даже геном клетки. По функциональным свойствам медиаторы ЦНС делятся на возбуждающие, тормозные и модулирующие. Возбуждающими медиаторами могут быть различные вещества, которые вызывают деполяризацию постсинаптической мембраны. Важнейшее значение имеют производные глутаминовой кислоты (глутамата), субстанция Р. Некоторые центральные нейроны имеют холинорецепторы, т.е. содержат на постсинаптической мембране рецепторы, которые реагируют с холинового соединениями, например, ацетилхолин в клетках Реншоу.. возбуждающими медиаторами могут быть также моноамины (норадреналин, дофамин, серотонин). € основания считать, что тип медиатора, который образуется в синапсе, обусловлен не только свойствами окончания, но и общим направлением биохимических процессов во всем нейроне. Природа тормозного медиатора до конца не установлена. Полагают, что в синапсах различных нервных структур эту функцию могут выполнять аминокислоты - глицин и ГАМК.
Электрические синапсы
Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока.
Нейрон - это структурно-функциональная единица нервной ткани. Это специализированная клетка, которая, наряду с общими физиологическими свойствами (возбудимость, проводимость), обладает и рядом специфических свойств :
- Воспринимать информацию - переводить информацию раздражителя на биологический язык клетки.
- Обрабатывать информацию - т.е. проводить анализ информации, синтез - соединение различных частей информации после анализа с получением нового качества.
- Кодировать информацию - превращать информацию в форму удобную для хранения в мозге.
- Формировать командный управляющий сигнал, который распространяется на другие клетки нейроны, мышечные клетки.
- Передача информации нейрона на другие структуры.
Нейроны способны контактировать с другими клетками и оказывать на них информационное воздействие (место контактов - синапс).
Все свои виды деятельности нейрон осуществляет за счѐт 3-х физиологических свойств (помимо возбудимости и проводимости):
Рецепция;
Электрогенез;
Нейросекреция.
В общем плане , все нейроны имеют тело – сому и отростки – дендриты и аксоны.
Их условно разделяют по структуре и функциям на следующие группы:
По форме тела: многоугольные, пирамидные, круглые, овальные.
По количеству и характеру отростков:
Униполярные – имеющие один отросток
Псевдоуниполярные – от тела отходит один отросток, который затем делится на 2 ветви.
Биполярные – 2 отростка, один дендритоподобный, другой аксон.
Мультиполярные – имеют 1 аксон и много дендритов.
По медиатору, выделяемому нейроном в синапсе: холинергические, адренергические, серотонинергические, пептидергические и т.д.
По функциям:
Афферентные, или чувствительные - служат для восприятия сигналов из внешней и внутренней среды и передачи их в ЦНС.
Вставочные , или интернейроны, промежуточные - беспечивают переработку, хранение и передачу информации к эфферентным нейронам. Их в ЦНС большинство.
Эфферентные или двигательные - формируют управляющие сигналы, и передают их к периферическим нейронам и исполнительным органам.
По физиологической роли: возбуждающие и тормозные.
Общими функциями нейронов ЦНС являются прием, кодирование, хранение информации и выработка нейромедиатора. Нейроны, с помощью многочисленных синапсов, получают сигналы в виде постсинаптических потенциалов. Затем перерабатывают эту информацию и формируют определенную ответную реакцию. Следовательно, они выполняют и интегративную, т.е. объединительную функцию .
Связь между нейронами , как видно, осуществляется через промежуток между концами аксона одного нейрона и дендритами другого. Если они лежат в достаточной близости, т. е. промежуток мал, то в этом месте может образоваться синаптический узел, или синапс, связывающий эти два нейрона.
Синапс
подобен сопротивлению в электрической цепи. Если это сопротивление велико, то связь между нейронами слабая и возбуждение одного нейрона не вызывает возбуждения другого. Если же «сопротивление» синапса мало, то имеется сильная связь и нейрон без труда возбуждается от аксона другого нейрона, с ним связанного.
Возбуждение нейрона происходит по принципу «все или ничего». Это значит, что нейрон может быть либо возбужден, и от клетки вдоль аксона к синаптическим узлам и далее к другим нейронам идет нервный импульс, либо не возбужден.
2.Гуморальная регуляция. Функции, механизмы взаимодействия гуморальных веществ с клетками мишенями. Место и роль желез внутренней секреции в регуляции функции.
Гуморальная регуляция - один из эволюционно ранних механизмов регуляции процессов жизнедеятельности в организме, осуществляемый через жидкие среды организма (кровь, лимфу, тканевую жидкость, полость рта) с помощью гормонов, выделяемых клетками, органами, тканями. У высокоразвитых животных, включая человека, гуморальная регуляция подчинена нервной регуляции и составляет совместно с ней единую систему нейрогуморальной регуляции. Продукты обмена веществ действуют не только непосредственно на эффекторные органы, но и на окончания чувствительных нервов (хеморецепторы) и нервные центры, вызывая гуморальным или рефлекторным путём те или иные реакции. Гуморальная передача нервных импульсов химическими веществами, медиаторами, осуществляется в центральной и периферической нервной системе. Наряду с гормонами важную роль в гуморальной регуляции играют продукты промежуточного обмена.
Биологическая активность жидких сред организма обусловлена соотношением содержания катехоламинов (адреналина и норадреналина, их предшественников и продуктов распада), ацетилхолина, гистамина, серотонина и других биогенных аминов, некоторых полипептидов и аминокислот, состоянием ферментных систем, присутствием активаторов и ингибиторов, содержанием ионов, микроэлементов и т. д.
В зависимости от строения гормона существуют два типа взаимодействия . Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют вторыми посредниками. Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет. Надежность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Посредники - это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Существует два главных способа передачи сигнала в клетки- мишени от сигнальных молекул с мембранным механизмом действия: аденилатциклазная (или гуанилатциклазная) системы; и фосфоинозитидный механизм.
Система циклазная – это система, состоящая из содержащихся в клетке аденозинциклофосфата, аденилатциклазы и фосфодиэстеразы, регулирующая проницаемость клеточных мембран, участвует в регуляции многих обменных процессов живой клетки, опосредует действие некоторых гормонов. То есть роль циклазной системы заключается в том, что они являются вторыми посредниками в механизме действия гормонов.
Система «аденилатциклаза - цАМФ». Мембраны фермент аденилатциклаза может находиться в двух формах - активированной и неактивированной. Активация аденилатциклазы происходит под влиянием гормон-рецепторного комплекса, образование которого приводит к связыванию гуанилового нуклеотида (ГТФ) с особым регуляторным стимулирующим белком (GS-белок), после чего GS-белок вызывает присоединение магния к аденилатциклазе и ее активацию. Так действуют активизирующие аденилатциклазу гормоны глюкагон, тиреотропин, паратирин, вазопрессин, гонадотропин и др. Некоторые гормоны, напротив, подавляют аденилатциклазу (соматостатин, ангиотензин-П и др.)
Под влиянием аденилатциклазы из АТФ синтезируется цАМФ, вызывающий активацию протеинкиназ в цитоплазме клетки, обеспечивающих фосфорилирование многочисленных внутриклеточных белков. Это изменяет проницаемость мембран, т.е. вызывает типичные для гормона метаболические и, соответственно, функциональные сдвиги. Внутриклеточные эффекты цАМФ проявляются также во влиянии на процессы пролиферации, дифференцировки, на доступность мембранных рецепторных белков молекулам гормонов.
Система «гуанилатциклаза - цГМФ». Активация мембранной гуанилатциклазы происходит не под непосредственным влиянием гормон-рецепторного комплекса, а опосредованно через ионизированный кальций и оксидантные системы мембран. Так реализуют свои эффекты натрийуретический гормон предсердий - атриопептид, тканевой гормон сосудистой стенки. В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны. Примерами могут служить стимуляция сокращений сердца под влиянием цАМФ и торможение их цГМФ, стимуляция сокращений гладких мышц кишечника цГМФ и подавление цАМФ.
Кроме аденилатциклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата.
Инозитолтрифосфат - это вещество, которое является производным сложного липида - инозитфосфатида. Оно образуется в результате действия специального фермента - фосфолипазы "С", который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора. Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30 % состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс "Са+2-кальмодулин" становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса "Са+2-кальмодулин" на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться.
Таким образом, в роли "вторых посредников" для передачи сигналов от гормонов в клетках-мишенях могут быть: циклические нуклеотиды (ц-АМФ и ц-ГМФ); ионы Са; комплекс "Са-кальмодулин"; диацилглицерин; инозитолтрифосфат.
Механизмы передачи информации от гормонов внутри клеток-мишеней с помощью перечисленных посредников имеют общие черты : одним из этапов передачи сигнала является фосфорилирование белков; прекращение активации происходит в результате специальных механизмов, инициируемых самими участниками процессов, - существуют механизмы отрицательной обратной связи.
Гормоны являются основными гуморальными регуляторами физиологических функций организма, и в настоящее время хорошо известны их свойства, процессы биосинтеза и механизмы действия. Гормоны являются высокоспецифичными веществами по отношению к клеткам-мишеням и обладают очень высокой биологической активностью.
Железы внутренней секреции – специализированные органы, не имеющие выводных протоков и выделяющие секрет в кровь, церебральную жидкость, лимфу через межклеточные щели.
Физиологическая роль желез внутренней секреции связана с их влиянием на механизмы регуляции и интеграции, адаптации, поддержания постоянства внутренней среды организма.
Эта клетка имеет сложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов.
Основные свойства нейрона - возбудимость и проводимость.
Возбудимость присуща клеткам всех тканей. Но у нервных клеток она очень высока. Раздражения вызывают в клетке ответную реакцию. Способность воспринимать раздражения и отвечать на них называется возбудимостью.
Раздражение вызывает в нейроне сложный процесс - возбуждение.
Оно мгновенно охватывает весь нейрон, а затем распространяется на все нервыные клетки, с которыми соприкасается этот нейрон. Способность нейрона передавать возбуждение называется проводимостью. Из центральной нервной системы к органам передают возбуждение центробежные нейроны.
От органона в центральную нервную систему оно проводится по центростремительным нейронам. Тела нейронов лежат преимущественно в центральной нервной системе. Они серого цвета и образуют серое вещество головного и спинного мозга.
Возбуждение проводится с различной скоростью:
от 0,5 до 120 м/сек. Быстрее всего оно передается к мышцам. Если в мышцу поступает поток нервных импульсов, то она сокращается.
При ходьбе мышцы ног попеременно сокращаются и расслабляются.
Расслабление мышц происходит под влиянием торможения. В том случае, когда синапсы задерживают нервные импульсы, развивается процесс торможения. Нервные импульсы не доходят до мышцы и она расслабляется.
Нейрон – это нервная клетка. Он является основной структурно-функциональной единицей нервной системы. Вместе с нейроглиальными клетками нейроны формируют нервную ткань, которая относится к возбудимым и характеризуется возбудимостью, проводимостью и лабильностью . Некоторые нейроны обладают автоматией .
в нейроне можно выделить три структурно-функциональных сегмента (рис. 1):
· рецепторный – в него входят дендриты и тело нейрона;
· передающий – это аксон нейрона на всем его протяжении до пресинаптических терминалей;
эффекторный – это пресинаптические терминали аксона
Классификация:
По локализации:
а) центральные – тела которых располагаются в ЦНС;
б) периферические – тела которых располагаются вне ЦНС (например, в спинальных или вегетативных ганглиях).
По функции:
а) афферентные (сенсорные) – несут информацию в ЦНС о состоянии внешней или внутренней среды. Они имеют высокочувствительное окончание – рецептор, в котором происходит трансформация энергии раздражителя в биоэлектрический сигнал;
б) эфферентные (моторные) – несут информацию от ЦНС к рабочему органу (мышце или секреторной клетке);
в) вставочные (ассоциативные, контактные, интернейроны) – обеспечивают связь между сенсорными и моторными нейронами. Они составляют 95-97% серого вещества головного и спинного мозга.
По физиологическому (функциональному) эффекту:
а) возбуждающие – передают возбуждение на последующую структуру;
б) тормозные – препятствуют процессу возбуждения на последующей структуре.
По функциональной (импульсной) активности:
а) нейроны с фоновой импульсной активностью . Они в состоянии покоя постоянно возбуждены и посылают импульсы на другие нейроны или на рабочий орган. Эти нейроны могут усиливать или ослаблять свою импульсную активность в зависимости от функционального состояния;
б) «молчащие» нейроны – они не имеют фоновой импульсной активности, но при действии раздражителя импульсация появляется и проявляется тем больше, чем больше функциональная активность нейрона.
В зависимости от количества модальностей раздражителя, адекватных для нейрона:
а) мономодальные – для них адекватна только одна модальность раздражителя;
б) полимодальные – для них адекватны две и более модальности
По медиатору, который выделяется в окончаниях аксона нейрона:
холинэргические, адренергические, серотонинэргические, пептидэргические, тауринэргические и др.
Кроме этого выделяют:
· релейные (проекционные) нейроны – это нейроны сенсорных путей в центральной части проводникового отдела анализатора. Они участвуют в проведении возбуждения к корковому отделу анализатора (см. в учебнике по нормальной физиологии раздел «Анализаторы»);
· нейросекреторные – отвечающие на нервный импульс секрецией гормонов (например, в гипоталамусе).
Связь с нейроглией
Эта связь крайне необходима для нормального функционирования нейрона. Роль нейроглии: клетки, входящие в нервную ткань и выполняющие важную функцию по обеспечению нормальной работы нейронов. Известно, что глиальных клеток больше, чем нервных. В детском возрасте их количество составляет 100 –140 млрд, а с возрастом увеличивается, так как глиальные клетки замещают погибшие нейроны.
Выделяют несколько функций нейроглии:
а) опорная: Глиальные клетки образуют основу (матрицу), на которой располагаются нейроны. Эту функцию в основном выполняют астроциты.
б) защитная: Нейроны окружены глиальными клетками, которые формируют более или менее выраженную оболочку вокруг тела и отростков, защищая их как от механических повреждений, так и от действия различных химических ве-ществ. Эту функцию в основном выполняют олигодендроциты.
Клетки микроглии обладают фагоцитарной активностью и разрушают как попавшие сюда микроорганизмы, так и погибшие в результате апоптоза или некроза нейроны.
в) трофическая: Эта функция заключается в том, что нейроны фактически не имеют связи с кровеносными сосудами и получают питательные вещества через глиальные клетки. Через них же нейрон выделяет во внутреннюю среду продукты метаболизма. Сигналом к усилению трофической функции является выход ионов калия из более активно работающего нейрона.
г) барьерная: Глиальные клетки защищают нейрон от действия токсических веществ, источником которых может быть как внешняя среда, так и нарушенный обмен веществ. Таким образом, глиальные клетки участвуют в формировании гематоэнцефалического барьера (ГЭБ).
д) секреторная: Глиальные клетки секретируют целый ряд биологически активных веществ, которые оказывают влияние на различные свойства и функции нейрона (возбудимость, проводимость, лабильность, метаболизм, двигательную активность и др.).
е) двигательная: Глиальные клетки «пульсируют» так же, как и нейроны, но частота этих пульсаций больше (до 20 в час). Эта активность глии способствует аксоплазматическому току жидкости в нейроне.
А ксоплазматический ток жидкости в нейроне:Это движение аксоплазмы по нейрофиламентам и микротубулам аксона от тела нейрона к его синаптическим терминалям (антероградный ток) и в обратном направлении (ретроградный ток).
По скорости различают быстрый и медленный аксоплазматический ток. Быстрый идет со скоростью 5-10 мм/час, а медленный 1-3 мм/сутки.
Антероградный аксоплазматический ток: Это движение аксоплазмы от тела нейрона к его синаптическим терминалям. Подобным образом доставляются к пресинаптическому окончанию ферменты, необходимые для синтеза медиатора, сам медиатор, нейротрофины – вещества, влияющие на метаболизм соседних нейронов, нейромодуляторы, влияющие на процесс передачи возбуждения в синапсе (см. вопрос 4ж).
Ретроградный ток аксоплазмы: Это движение аксоплазмы в противоположном направлении от синаптических терминалей аксона к его телу. Таким образом возвращаются продукты метаболизма, продукты, появившиеся в результате распада медиатора, нейротрофины и др.
Каковы показатели функциональной активности нейрона?
Таких показателей достаточно много и все они могут быть объединены в три группы: структурные, биохимические, физиологические.
Структурные:
а) уменьшение хроматофильной субстанции (телец Ниссля);
б) увеличение размеров ядра;
в) увеличение количества митохондрий;
г) увеличение количества «шипиков» на теле нейрона, то есть увеличение количества синаптических контактов нейрона.
Биохимические:
а) усиление обмена белков, жиров и углеводов;
б) усиление потребления кислорода;
в) увеличение активности ферментов.
Физиологические:
а) увеличение двигательной активности нейрона и нейроглии;
Нейрон. Его свойства и функции Факультет психологии П С Д – Д С- 2 Филиппова Ольга Москва Нейрон (от др.-греч. νεῦρον - волокно, нерв) - это структурно-функциональная единица нервной системы. Эта клетка имеет сложное строение, высоко специализирована и по структуре содержит ядро, тело клетки и отростки. В организме человека насчитывается более ста миллиардов нейронов. Сложность и многообразие функций нервной системы определяются взаимодействием Между нейронами, которое, в свою очередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами или мышцами и железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих Электрический заряд, который движется вдоль нейрона. Строение Тело клетки тело Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), снаружи ограничена мембраной из двойного слоя липидов(билипидный слой). Липиды состоят из гидрофильных головок и гидрофобных хвостов, расположены гидрофобными хвостами друг к другу, образуя гидрофобный слой, который пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране Находятся белки: на поверхности (в форме глобул), на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы. Нейрон состоит из тела диаметром от 3 до 130 мкм, содержащего ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также изотростков. Выделяют два вида отростков:дендриты и аксоны. Нейрон имеет развитый и сложный цитоскелет, проникающий в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и дендрит упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибриллразного диаметра: аксон Микротрубочки (Д = 20-30 нм) - состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) - вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) - состоят из белков актина и миозина, особенно выражены в растущих нервных отростках и в нейроглии. В теле нейрона выявляется развитый синтетический аппарат, гранулярная ЭПС н ейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Аксоны и дендриты Аксон - обычно длинный отросток, приспособленный для проведения возбуждения от тела нейрона. Дендриты - как правило, короткие и сильно разветвлённые отростки, служащие главным местом образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов). Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20-и тысяч) другими нейронами. аксон дендриты - Дендриты делятся дихотомически, - аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии. - Дендриты не имеют миелиновой оболочки, - аксоны же могут её иметь. Местом генерации возбуждения у большинства Нейронов является аксонный холмик - образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной. Синапс Си́ напс (греч. σύναψις, от συνάπτειν - обнимать, обхватывать, пожимать руку) - место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсывызывают деполяризацию нейрона, другие - гиперполяризацию; первые являются возбуждающими, вторые - тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов. Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Классификация Структурная классификация Функциональная классификация Структурная классификация Безаксонные нейроны Униполярные нейроны Биполярные нейроны Мультиполярные нейроны Псевдоуниполярные нейроны Функциональная классификация Афферентные нейроны Эфферентные нейроны Ассоциативные нейроны Секреторные нейроны Структурная классификация Безаксонные нейроны - небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено. Униполярные нейроны - нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге. Биполярные нейроны - нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах - сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях. Мультиполярные нейроны - нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе. Псевдоуниполярные нейроны - являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях. Функциональная классификация Афферентные нейроны (чувствительный, сенсорный или рецепторный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания. Эфферентные нейроны (эффекторный, двигательный или моторный). К нейронам данного типа относятся конечные нейроны - ультиматные и предпоследние - не ультиматные. Ассоциативные нейроны (вставочные или интернейроны) - группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные. Секреторные нейроны - нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами. Развитие и рост нейрона Нейрон развивается из небольшой клетки предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, имеющиеся в теле нейрона Основные свойства нейронов Раздражимость - способность нервной клетки отвечать на различные раздражения биохимическими изменениями, сопровождающимися нарушением ионного равновесия и деполяризацией электрических зарядов на мембранах клетки вместе раздражения. Раздражимость присуща всем клеткам, и особенно нервным, связанным с чувствительным восприятием запаховых, звуковых, световых и других раздражителей. Раздражимость - пусковой механизм проявления другого свойства - возбудимости. Возбудимость - способность отдельных частей нервной клетки генерировать электрохимические импульсы, т. е. отвечать на раздражение возбуждением. Для перехода нервной клетки в состояние возбуждения необходимо, чтобы сила действующего раздражителя достигла критического предела - пороговой величины. Способность нейрона отвечать возбуждением на наименьшую силу раздражителя называется нижним порогом возбудимости. Чем чувствительнее нервная клетка к раздражению, тем меньше порог возбудимости, и, следовательно, даже самый слабый раздражитель может вызвать возбуждение. Величина возбуждения нейрона зависит от силы раздражителя и возрастает по закону силовых отношений до определенного предела - верхнего порога возбудимости. Применение раздражителей сверхпороговой силы создает в нейроне запредельное торможение, которое охраняет нервную клетку от перевозбуждения Одиночное раздражение обычно вызывает серию импульсов определенной силы, продолжительности и частоты. В разных нервных клетках частота импульсов различная - от 100 до 1000 в секунду. Сила и продолжительность импульсов возбуждения зависит от характера раздражения. Проводимость - способность нейрона проводить импульсы возбуждения с определенной скоростью, в неизменном ритме и силе. Возбуждение по нервному волокну может распространяться в обе стороны от раздражаемого участка. В разных нервных клетках скорость проведения возбуждения неодинакова и зависит от физиологического состояния нейрона и толщины волокна. В чувствительных нейронах возбуждение распространяется со скоростью 100–120 метров в секунду, в двигательных - 60–100, а в вегетативной нервной системе - 5–7. Лабильность (подвижность) - способность нервной клетки принимать и передавать максимальное число импульсов за единицу времени без искажения. Подвижность двигательных нейронов не более 500 импульсов в секунду. Лабильность обеспечивает направленное распределение и проведение импульсов возбуждения нужной частоты по определенным нервным путям. В процессе роста и развития организма, а также при систематической тренировке, лабильность увеличивается и обеспечивает динамичность нервной системы, при утомлении и старении - уменьшается. Инертность - способность нервной клетки накапливать и хранить в себе следы возбуждения и торможения. Полученная информация откладывается в дендритах, соме клетки, хромосомах ядра в виде биохимических изменений ДНК и РНК плазмы. Это свойство нейронов обеспечивает память организма, которая имеет решающее значение в процессе обучения животных. Утомляемость - естественный процесс снижения работоспособности клетки при длительном возбуждении или торможении. Проявляется в виде уменьшения силы возбуждения, замедления частоты ритма импульсов и скорости их проведения. Отдых нервных клеток или смена нервной деятельности снимает утомление, и все свойства восстанавливаются. Торможение - процесс, обратный возбуждению. Заключается в ослаблении, остановке или предупреждении возникновения возбуждения. Торможение - активный процесс, распространяясь по нервным клеткам, он обеспечивает согласованную работу отдельных органов и всего организма в целом. Регенерация - способность нервной клетки восстанавливать утраченные или поврежденные отростки путем прорастания. Нервные клетки не размножаются, погибшие нейроны не восстанавливаются. Волокна нервной клетки способны прорастать, если сохранилось тело клетки. Основные функции нейронов Рецепторная функция обеспечивает восприятие определенных раздражителей из внешней и внутренней среды организма. Рецепторные клетки - это видоизмененные нейроны, воспринимающие определенный вид энергии Поступающее из внешней или внутренней среды. Рецепторы, воспринимающие раздражения из внешней средой называют экстерорецепторами, из внутренней среды - интерорецепторами. Сенсорная функция чувствительных нейронов обеспечивает анализ воспринятых раздражений, формирование определенных ощущений и четкую дифференцировку многочисленных раздражителей, воздействующих из внешней и внутренней среды. Информационная функция промежуточных нейронов обеспечивает накопление, сохранение и выдачу информации, поступившей из внешней и внутренней среды. Информация в нейронах кодируется как память и в нужных случаях выдается в виде слабых импульсов возбуждения. Моторная функция двигательных нейронов обеспечивает формирование и передачу импульсов возбуждения определенной силы и частоты к соответствующим органам движения или другим исполнительным органам и тканям. Таким образом, основными функциями нейронов являются: восприятие раздражений, их переработка и передача нервных возбуждений на другие нейроны или рабочие органы. Через нейроны осуществляется передача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участками тела и органами. В нейронах происходят сложнейшие процессы обработки и запоминания информации. С помощью нейронов формируются рефлексы. Список литературы Немечек С. и др. Введение в нейробиологию, Avicennum: Прага, 1978, 400 c. Физиология человека под редакцией В.М.Покровского, Г.Ф.Коротько Анисимов В.Н. - Молекулярные и физиологические механизмы старения
Последние материалы сайта
Симптомы

Карта-схема комплексного обследования
Логопедическое обследование детей с церебральным параличом Логопедическое обследование детей, страдающих церебральным параличом, строится на общем системном подходе, который опирается на представление о речи как о сложной функциональной системе, структ
Методы диагностики

Пять компотов из вишни на зиму
Александр Гущин За вкус не ручаюсь, а горячо будет:) Содержание Яркая, красивая ягода вишня содержит в себе огромное количество полезных микроэлементов и витаминов. Ее универсальность в кулинарии позволяет приготовить массу хороших, вкусных, уникаль
Болевые синдромы

Готовим изумительную пшенную кашу на молоке в мультиварке редмонд Как сварить пшенную кашу скороварке
Ни одна кастрюля в мире не сварит пшенную кашу на молоке так вкусно, как это сделает мультиварка. И ни одна плита не справится с ее приготовлением так быстро, как мультиварка-скороварка. Простой и здоровый завтрак – сладкая пшенная каша на молоке, сдобрен
Синдромы

Рыцарь монет таро значение в отношениях
Название: Рыцарь пентаклей, Рыцарь денариев, Рыцарь монет, Воин монет, Рыцарь бубен, Путешественник, Завоеватель Дел, Всадник, Кавалер, Полезный человек, Повелитель Земель Диких и Плодородных. Значение по Папюсу: Выгода, Прибыль, Польза, Интерес. Важное
Методы лечения

Гороскоп на август телец девушка
Томный август у многих Тельцов станет благоприятным временем для развития личных отношений. В этот период у представителей данного знака Зодиака возникает возможность использовать на полную катушку свое природное обаяние для успеха в сердечных делах. Чтоб
Анатомия и физиология

История россии с древнейших времен до наших дней
Историк А.Н.Сахаров – доктор? Чего? /памфлет/ Вы, конечно, помните "Суды времени" на 5-канале петербургского ТВ, проводимые двумя либерал-поборниками Н.Сванидзе и Л.Млечиным при участии со стороны защиты С.Кургиняна. В определенном месте и назначенный ча