Поперечно полосатая скелетная мускулатура. Мышечная ткань. Поперечно-полосатая мышечная ткань
Здесь описаны основные принципы строения мышечной ткани. Рассматриваются следующие темы.
- Строение поперечнополосатых мышц (мышечные волокна, соединительнотканные фасции, кровоснабжение и иннервация мышц).
- Рецепторы мышечной ткани.
- Строение мышечно-сухожильного соединения, гистологическое строение сухожилий и их прикрепления к кости (костносухожильное соединение).
- Описание различных типов мышц и мышечных волокон.
- Функциональное значение поперечнополосатых мышц.
- Анатомия важнейших мышц: начало, прикрепление, иннервация, функция и значение в различных видах спорта.
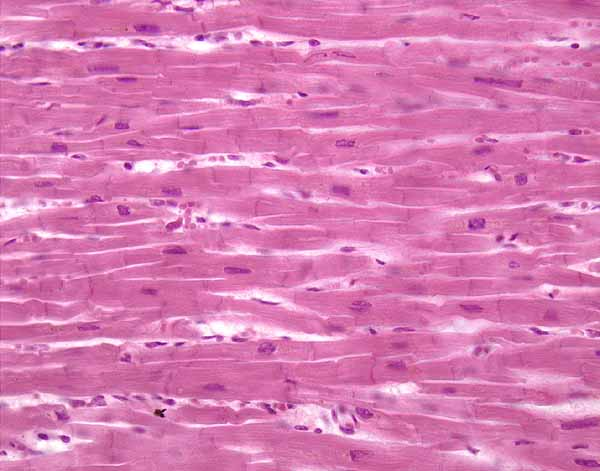
Мышцы позволяют организму человека совершать различные движения. В цитоплазме мышечных волокон (клеток мышечной ткани) находится большое количество специальных белков (актомиозина), благодаря которым возможно мышечное сокращение. В организме человека выделяют три вида мышечной ткани, различающихся но своим морфологическим и физиологическим свойствам.
- Поперечнополосатая, или скелетная, мышечная ткань — состоит из мышечных волокон цилиндрической формы и иннервируется соматической нервной системой (произвольная мускулатура).
- Гладкая мышечная ткань — состоит преимущественно из веретенообразных клеток. Гладкая мышечная ткань находится в стенках внутренних органов и кровеносных сосудов, а также в корнях волос, железах внешней секреции и глазном яблоке (Tillmann, 1998). Гладкая мышечная ткань получает иннервацию от вегетативной нервной системы (непроизвольная мускулатура) (Silbernagl, Despopoulos, 1983). Некоторые гладкомышечные волокна получают автономную иннервацию от клеток — водителей ритма через щелевидные контакты (нексусы).
- Поперечнополосатая сердечная мышечная ткань — состоит из поперечно исчерченных кардиомиоцитов, расположенных параллельно друг другу и соединенных так называемыми вставочными дисками. Сердечная мышечная ткань получает импульсы от автономных клеток — водителей ритма» на нее также оказывает регулирующее влияние вегетативная нервная система (Mauer, 2006).
Тело человека имеет 430 мышц, которые составляют 40-50% его массы и, таким образом, являются самой распространенной тканью человека (Cabri, 1999). Скелетные мышцы прикрепляются к костям скелета с помощью сухожилий, причем прикрепление мышц может быть прямым или непрямым. Мышечная ткань вместе со вспомогательными структурами (соединительнотканные оболочки — фасции, кровеносные сосуды, нервы, синовиальные сумки, влагалища сухожилий, нервно-мышечные веретена и рецепторы сухожилий) образует эффективную систему, гармонично передающую силу на опорно-двигательный аппарат. Благодаря своему строению скелетная мускулатура, с одной стороны, обеспечивает движения, а с другой — участвует в поддержании позы. При этом мышечная система выполняет и защитную функцию при воздействии внешних сил.
- Мышечное волокно не является клеткой. Оно состоит из мио-симпласта и миосателлитоцитов (клеток-спутниц), покрытых общей базальной мембраной. Миосимпласт представляет собой совокупность слившихся клеток, в нем имеется большое количество ядер, расположенных по периферии мышечного волокна. Мышечное волокно является структурной единицей мышечной ткани.
- Актомиозин состоит из белков актина и миозина, которые образуют актомиозиновый комплекс, обладающий АТФазной активностью, т.е. способностью расщеплять АТФ, при этом освобождается энергия, необходимая для обеспечения сокращений мышц.
Введение
Мышцы, органы тела животных и человека, за счет сокращения и расслабления которых происходят все движения тела и внутренних органов. Мышцы образуются мышечной тканью в сочетании с другими тканевыми структурами - соединительнотканными компонентами, нервами и кровеносными сосудами.
Мышечная ткань состоит из мышечных клеток, которым в наибольшей степени присуще свойство сократимости. Издавна эти клетки называют мышечными волокнами; данный термин указывает лишь на то, что мышечные клетки имеют вытянутую форму. Сократимость (одно из основных свойств животных клеток, которое наиболее развито в мышечных тканях) мышечных волокон обеспечивается аппаратом, образованным сократительными белками (актином и миозином), взаимодействие которых, протекающее с использованием энергии (АТФ), приводит к сокращению клеток (укорочению). Вслед за сокращением наступает расслабление, и тогда они возвращаются к своей исходной длине. Благодаря такому свойству клеток мышечной ткани достигается все многообразие двигательных функций организма и протекающих в нем механических процессов.
Существует три типа мышечной ткани и соответственно мышц, различающихся по структуре мышечных волокон и характеру иннервации:
1. Скелетная (поперечно-полосатая) мышечная ткань
2. Сердечная поперечно - полосатой мышечная ткань
3. Гладкая мышечная ткань
Скелетная (поперечно-полосатая) мышечная ткань -- упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов; один из типов мышечной ткани. Образует скелетную мускулатуру человека и животных, предназначенную для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания.
Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) диаметром от 50 до 100 мкм. Клетки многоядерные, содержат до 100 и более ядер. Микроскопическое изучение показало, что волокно скелетной мышцы по всей длине имеет регулярную поперечную исчерченность в виде чередующихся светлых и темных участков, (поперечнополосатая мышечная ткань образована мышечными клетками, содержащими миофибриллы, которые состоят из миозиновых и актиновых протофибрилл, взаимное положение которых создает поперечную исчерченность),что послужило основанием для другого названия - поперечнополосатые мышцы.
Функции скелетных мышц находятся под контролем центральной нервной системы, т.е. контролируются нашей волей, поэтому их называют также произвольными мышцами. Однако они могут находиться в состоянии частичного сокращения и независимо от нашего сознания; такое состояние называют тонусом. мышечный ткань волокно
Сердечная поперечно - полосатая мышечная ткань
Структурно-функциональной единицей сердечной поперечно - полосатой мышечной ткани является клетка - кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
Типичные или сократительные кардиомиоциты, образующие своей совокупностью миокард;
Атипичные кардиомиоциты, составляющие проводящую систему сердца и подразделяющиеся в свою очередь на три разновидности.
Сократительный кардиомиоцит представляет собой почти прямоугольную клетку 50-120 мкм в длину, шириной 15-20 мкм, в центре которой локализуется обычно одно ядро. Покрыт снаружи базальной пластинкой. В саркоплазме кардиомиоцита по периферии от ядра располагаются миофибриллы, а между ними и около ядра локализуются в большом количестве митохондрии. В отличие от скелетной мышечной ткани, миофибриллы кардиомиоцитов представляют собой не отдельные цилиндрические образования, а по существу сеть, состоящую из анастомозирующих миофибрилл, так как некоторые миофиламенты как бы отщепляются от одной миофибриллы и наискось продолжаются в другую. Кроме того, темные и светлые диски соседних миофибрилл не всегда располагаются на одном уровне, и потому поперечная исчерченность в кардиомиоцитах выражена не столь отчетливо, как в скелетных мышечных волокнах. Саркоплазматическая сеть, охватывающая миофибриллы, представлена расширенными анастомозирующими канальцами. Терминальные цистерны и триады отсутствуют. Т-канальцы имеются, но они короткие, широкие и образованы не только углублением плазмолеммы, но и базальной пластинки. Механизм сокращения в кардиомиоцитах практически не отличается от такового в скелетных мышечных волокнах.
Сократительные кардиомиоциты, соединяясь встык друг с другом, образуют функциональные мышечные волокна, между которыми имеются многочисленные анастомозы. Благодаря этому из отдельных кардиомиоцитов формируется сеть - функциональный синтиций. Наличие щелевидных контактов между кардиомиоцитами обеспечивает одновременное и содружественное их сокращение вначале в предсердиях, а затем и в желудочках.
Области контактов соседних кардиомиоцитов носят название вставочных дисков. Фактически, никаких дополнительных структур (дисков) между кардиомиоцитами нет. Вставочные диски - это места контактов цитолеммы соседних кардиомиоцитов, включающие в себя простые, десмосомные и щелевидные контакты. Обычно во вставочных дисках различают поперечный и продольный фрагменты. В области поперечных фрагментов имеются расширенные десмосомные соединения. В этих же местах с внутренней стороны плазмолемм прикрепляются актиновые филаменты саркомеров. В области продольных фрагментов локализуются щелевидные контакты. Посредством вставочных дисков обеспечивается как механическая, так и метаболическая (прежде всего ионная) связь кардиомиоцитов.
Сократительные кардиомиоциты предсердий и желудочков несколько отличаются между собой по морфологии и функциям. Так, кардиомиоциты предсердий в саркоплазме содержат меньше миофибрилл и митохондрий, в них почти не выражены Т-канальцы, а вместо них под плазмолеммой выявляются в большом числе везикулы и кавеолы - аналоги Т-канальцев. Кроме того, в саркоплазме предсердных кардиомиоцитов у полюсов ядер локализуются специфические предсердные гранулы, состоящие из гликопротеиновых комплексов. Выделяясь из кардиомиоцитов в кровь предсердий, эти вещества влияют на уровень давления крови в сердце и сосудах, а также препятствуют образованию тромбов в предсердиях. Следовательно, предсердные кардиомиоциты, кроме сократительной, обладают и секреторной функцией. В желудочковых кардиомиоцитах более выражены сократительные элементы, а секреторные гранулы отсутствуют.
Вторая разновидность кардиомиоцитов - атипичные кардиомиоциты образуют проводящую систему сердца, состоящую из:
Синусо-предсердный узел;
Предсердно-желудочковый узел;
Предсердно-желудочковый пучок (пучок Гиса), ствол, правую и левую ножки;
Концевые разветвления ножек - волокна Пуркинье.
Атипичные кардиомиоциты обеспечивают генерирование биопотенциалов, их проведение и передачу на сократительные кардиомиоциты.
По своей морфологии атипичные кардиомиоциты отличаются от типичных рядом особенностей:
Они крупнее (длина 100 мкм, толщина 50 мкм);
В цитоплазме содержимся мало миофибрилл, которые расположены неупорядочено и потому атипичные кардиомиоциты не имеют поперечной исчерченности;
Плазмолемма не образует Т-канальцев;
Во вставочных дисках между этими клетками отсутствуют десмосомы и щелевидные контакты.
Атипичные кардиомиоциты различных отделов проводящей системы отличаются между собой по структуре и функциям и подразделяются на три основные разновидности:
· Р-клетки (пейсмекеры) - водители ритма (I типа);
· переходные клетки (II типа);
· клетки пучка Гиса и волокон Пуркинье (III тип).
Клетки I типа (Р-клетки) составляют основу синусо-предсердного узла, а также в небольшом количестве содержатся в атриовентрикулярном узле. Эти клетки способны самостоятельно генерировать с определенной частотой биопотенциалы и передавать их на переходные клетки (II типа), а последние передают импульсы на клетки III типа, от которых биопотенциалы передаются на сократительные кардиомиоциты.
Источники развития кардиомиоцитов - миоэпителиальные пластинки, представляющие собой определенные участки висцеральных листков спланхнотома, а конкретнее из целомического эпителия этих участков.
Мышечные клетки обычно называют мышечными волокнами, потому что они постоянно вытянуты в одном направлении. Сократимость у мышечных клеток достигла наибольшего развития: они состоят из сокращающихся элементов, называемых миофибриллами, расположенных вдоль оси длинной волокна и придающих ему продольную исчерченность. Это характерно для всех видов мышечной ткани. Миофибриллы некоторых типов мышечной ткани состоят из перемежающихся плотноуложенных темных и светлых полос. В результате волокно имеет поперечную исчерченность и называется поперечно-полосатым мышечным волокном.
Классификация мышечных тканей проводится на основании строения ткани (гистологически): по наличию или отсутствию поперечной исчерченности, и на основании механизма сокращения - произвольного (как в скелетной мышце) или непроизвольного (гладкая или сердечная мышцы), т.е. по физиологическому признаку. Плазматическая мембрана мышечного волокна называется сарколеммой, и в зависимости от типа мышечной ткани она имеет различную степень развития.
Цитоплазма клеток мышечной ткани называется саркоплазмой, во всех трех видах мышечной ткани цитоплазма состоит из сократительных белков - актина и миозина и богата митохондриями, содержащими ферменты для активного метаболизма и сократительных движений мышечного волокна. Скелетная и сердечная мышечные ткани изобилуют гладким эндоплазматическим ретикулумом (саркоплазматическим реткулумом).
Классификация мышечной ткани.
А) По гистологическому признаку:
Неисчерченная:
Гладкая мышечная ткань
Исчерченная:
Сердечная мышца
Б) По физиологическому признаку:
Непроизвольная:
Гладкая мышечная ткань
Сердечная мышца
Произвольная:
Поперечно-полосатая мышечная ткань
Гладко-мышечная ткань.
Из гладкой мышечной ткани построены стенки висцеральных органов, мышечные оболочки кровеносных сосудов (артерий, артериол, вен, кроме венул). Длина мышечных волокон их колеблется от 20 до 500 мкм, а ширина в месте расположения ядра составляет около 5 мкм, в направлении обоих концов волокно истончается и заостряется. Ядро на поперечном срезе часто не визуализируется. Если же оно видно, то форма его сферическая. На продольном срезе ядро имеет овальную форму: при сокращении волокна оно может стать штопорообразным.
Сарколемму гладкой мышечной ткани очень трудно выявить при помощи световой микроскопии. Гладкие мышечные волокна окружены каркасом ретикулиновых волокон, погруженных в ШИК-положительное основное вещество. Считается, что ретикулиновые волокна служат ограничением для мышечного волокна, удерживая их при сокращении. При исследовании под электронным микроскопом выявляется, что с наружной стороны сарколемма покрыта базальной пластинкой. Гладкомышечные клетки могут увеличиваться в количестве с помошью митозов (гиперплазия), а также в размере (гипертрофия). Обычно гладкомышечные волокна располагаются в несколько слоев, в которых широкая часть одного волокна примыкает к суженным, вытянутым краям других волокон.
Гладкая мускулатура иннервируется вегетативной нервной системой: как симпатической, так и парасимпатической ее частей (стенка кишечника). В других случаях случаях сокращения мышечных органов осуществляется исключительно симпатической системой (стенка артерии), или парасимпатической (сфинктер зрачка). Нервные окончания могут располагаться только на поверхностных клетках слоя волокон: импульс проходит через слой от клетки к клетке через щелевые контакты (нексусы), области сужения межклеточного пространства с низким электрическим сопротивлением. Однако, в некоторых случаях, иннервируется каждая гладкомышечная клетка (радужная оболочка глаза и семявыносящий проток). В цитоплазме гладкомышечных клеток с помощью электронной микроскопии можно увидеть митохондрии, аппарат Гольджи, гликоген, шероховатый эндоплазматический ретикулум и рибосомы; эти органеллы располагаются у полюсов ядра. Цитоплазма также содержит интенсивно окрашенные, сигарообразные элементы, содержащие альфа-актин, объединенные в систему постоянных цитоплазматических филаментов промежуточного диаметра (10 нм) между тонкими актиновыми филаментами (7 нм толщиной) и толстыми миозиновыми филаментами (17 нм тощиной), которые также встречаются здесь. Сарколемма видна в местах инвагинации в продольные ряды пузырьков (кавеолы), которые тесно связаны с трубочками саркоплазматического ретикулума. Они регулируют концентрацию катионов кальция в клетке и проводят сократительные импульсы внутрь клетки (подобно Т-трубочкам поперечнополосатой мышечной ткани). В цитоплазме также встречаются микротрубочки, состоящие из тубулина.
Поперечно-полосатая мышечная ткань.
Волокна поперечно-полосатых мышц представляют собой многоядерные гигантские клетки цилиндрической формы: их длина колеблется от нескольких милиметров до нескольких сантиметров. Их толщина составляет от 10 до 150 мкм. Сарколемма достаточно развита, вдоль нее раположены многочисленные ядра, неравномерно разбросанные в пределах волокна. Они имеют овальную форму на продольном срезе и округлую на поперечном. Волокна характеризуются как продольной, так и поперечной исчерченностью, которая выражена значительнее. Наличие миофибрилл, идущих вдоль волокна, обусловливает продольную исчерченность. Миофибриллы состоят из чередующихся светлых изотропных и темных анизотропных полос; светлые называются А-полосами, а темные - I-полосами. Эти полосы плотно уложены, что придает мышечному волокну вид поперечной исчерченности. Окончания волокон тупо закруглены.
Любая поперечно-полосатая мышца, например двуглавые мышцы, одета в покров из соединительной ткани, состоящий из трех компонентов. Эпимизий из соединительной ткани, содержащей большое количество жировых клеток, покрывает мышцу (влагалище мышцы) и, погружаясь, разделяет брюшко мышцы на пучки. Каждый пучок волокон в пределах мышцы окружен толстым слоем соединительной ткани - перимизием. В пределах пучка каждое поперечно-полосатое мышечное волокно окружено тонкой прослойкой, богатой кровеносными сосудами - эндомизием.
С помощью электронной микроскопии установлено, что волокна поперечно- полосатых мышц имеют сложное строение. Миофибриллы состоят из двух типов, смещающихся относительно друг друга, миофиламентов: тонких актин-содержащих и толстых миозин-содеращих. I-полосы состоят из тонких миофиламентов, а А-полосы - из обоих типов. Точно по центру каждой I-полосы расположена темная Z-линия, к каждой стороне которой прикрепляются актин-содержащие тонкие миофиламенты. Расстояние между каждой парой Z-линий называется саркомером, элементарная сократимая единица поперечно-полосатых мышц. Тонкие миофиламенты выступают в область А-полосы, причем один конец миофиламента свободен и находится между толстыми миофиламентами, что способствует возникновению светлой Н-зоны в середине А-полосы. Толстые миофиламенты тянутся через всю А-полосу и их концы свободны. Миофибриллы расположены в строго определенном порядке: каждый толстый миофиламент окружен шестью равноудаленными тонкими миофиламентами, которые расположены в форме шестиугольника.
При сокращении, длина миофиламентов не уменьшается, а увеличивается только лишь их степень смещения относительно друг друга. В результате этого, I-полоса уменьшается и Н-зона тоже становится очень узкой, однако длина А-полосы практически не изменяется. На поперечном срезе миофибрилла имеет 3 вида, в зависимости от уровня среза: только на уровне тонких миофиламентов, толстых миофиламентов или на уровне двух типов филаментов одновременно. Две эллиптические митохондрии окружают каждую миофибриллу на уровне I-полосы.
При исследовании под электронным микроскопом удается выяснить, что сарколемма состоит из плазматической мембраны и вместе с хорошо развитой базальной пластинкой и сетью тонких ретикулиновых волокон. Плазматическая мембрана вдавливается и проходит через волокно на уровне границы между А- и I-полосами в виде вытянутой трубочки (Т-трубочка). При сокращении волна деполяризации распространяется вдоль сарколеммы и благодаря Т-трубочкам более или менее синхронно достигает всех частей миофибриллы. Саркоплазматический гладкий эндоплазматический ретикулум опоясывает каждую миофибриллу несколько раз таким образом, что по бокам каждой Т-трубочки параллельно располагаются две терминальные цистерны. Эти структуры, видимые рядом в поперечном разрезе, называются триадой. Трубочки и цистерны обеспечивают изоляцию зоны катионов кальция, что происходит во время расслабления мышцы. Большинство мышц человека содержит три типа поперечно-полосатых волокон: красные, белые и промежуточные. Преобладающим типом являются красные волокна, которые характеризуются малой толщиной и обилием пигментированного белка - миоглобина, и относительно небольшим количеством миофибрилл. Красные, или слабо сокращающиеся волокна, выполняют повторяющиеся сократительные движения и характерны для мышц туловища, ответственных за поддержание позы. Белые волокна толще и содержат большее количество миофибрилл, но меньше миоглобина, чем красные волокна. Они способны сокращаться быстрее и сравнительно быстро устают, поэтому они больше подходят для коротких вспышек активности. Промежуточные волокна по размерам и характеристикам занимают промежуточное положение между красными и белыми волокнами.
Иннервация поперечно-полосатых мышц.
К каждому экстрафузальному поперечно-полосатому мышечному волокну подходит окончание аксона от двигательного альфа-мотонейрона в переднем роге спинного мозга и двигательных ядрах черепно-мозговых нервов. Некоторые альфа- мотонейроны иннервируют полдюжины поперечно-полосатых мышечных волокон, как в случае наружных глазных мышц, где необходим тонкий контроль. Другие альфа- мотонейроны могут иннервировать до 500 мышечных волокон в большой мышце. Один мотонейрон вместе с иннервированными им мышечными волокнами образует двигательную единицу. Нервно-мышечное (мионевральное) соединение или двигательная концевая пластинка обычно располагается в середине волокна, здесь оно утолщено и имеет локальное скопление ядер и митохондрий. Миелиновая оболочка аксона исчезает и он разветвляется на несколько оголенных грушевидных окончаний, которые погружены в углубления на поверхности мышечного волокна - это называется синаптической щелью. Здесь сарколемма очень складчата (соединительные складки или субнейральные щели). Лишь аморфный компонент сарколеммы (базальная пластинка) разделяет плазматические мембраны аксона и мышечного волокна и он содержит фермент ацетилхолинэстеразу. Окончание аксона содержит много митохондрий и скопления синаптических пузырьков, в которых находится нейромедиатор - ацетилхолин.
Мышечные веретена представляют собой инкапсулированные скопления истонченных мышечных волокон, лишь частично содержащих небольшое количество миофибрилл; данные волокна называются интрафузальными в отличие от экстрафузальных волокон мышц в большинстве случаев. Функционально, они являются проприоцептивными, действующими в качестве рецепторов растяжения и имеющими двойную иннервацию. Интрафузальные волокна иннервируются гамма-мотонейронами, которые имеют окончания в двигательных концевых пластинках. Данные волокна также имеют анулоспиральные сенсорные нервные, или первичные, окончания вокруг их центрального участка.
Ткань - это совокупность схожих по строению клеток, которые объединены общими функциями. Практически все многоклеточные организмы состоят из разных типов тканей.
Классификация
У животных и человека в организме присутствуют следующие типы тканей:
- эпителиальная;
- нервная;
- соединительная;
- мышечная.
Эти группы объединяют по несколько разновидностей. Так, бывает жировой, хрящевой, костной. Также сюда относятся кровь и лимфа. Эпителиальная ткань существует многослойная и однослойная, в зависимости от строения клеток можно выделить также плоский, кубический, цилиндрический эпителий и т. д. Нервная бывает только одного вида. А о мышечном типе ткани мы поговорим подробнее в этой статье.
Виды мышечной ткани
В организме всех животных выделяют три ее разновидности:
- поперечно-полосатые мышцы;
- сердечная мышечная ткань.
Еще одним общим признаком всех видов мышц является то, что они способны сокращаться, однако у разных видов этот процесс происходит индивидуально.
Особенности миоцитов
Клетки гладкой мышечной ткани, как и поперечно-полосатой и сердечной, обладают вытянутой формой. Кроме того, в них есть особые органоиды, которые называются миофибриллы, или миофиламенты. В них содержатся сократительные белки (актин, миозин). Они необходимы для того, чтобы обеспечить движение мышцы. Обязательным условием функционирования мускула, кроме наличия сократительных белков, также является присутствие в клетках ионов кальция. Поэтому недостаточное или избыточное употребление продуктов с высоким содержанием данного элемента может привести к некорректной работе мускулатуры - как гладкой, так и поперечно-полосатой.
Кроме того, в клетках присутствует еще один специфический белок - миоглобин. Он необходим для того, чтобы связываться с кислородом и запасать его.
Что касается органоидов, то кроме наличия миофибрилл особенным для мышечных тканей является содержание большого количества в клетке митохондрий - двумембранных органоидов, отвечающих за И это неудивительно, так как мышечному волокну для сокращения необходимо большое количество энергии, вырабатываемой при дыхании митохондриями.
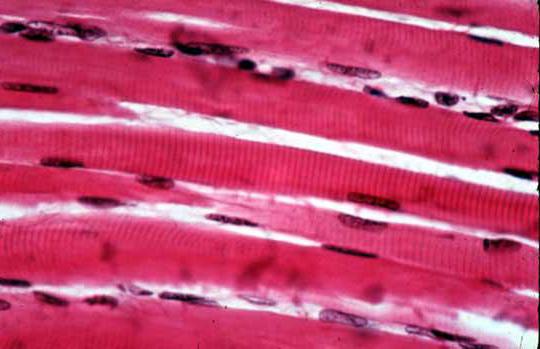
В некоторых миоцитах также присутствует более чем одно ядро. Это характерно для поперечно-полосатой мускулатуры, в клетках которой может содержаться около двадцати ядер, а иногда эта цифра доходит и до ста. Это связано с тем, что волокно поперечно-полосатой мышцы сформировано из нескольких клеток, объединенных впоследствии в одну.
Строение поперечно-полосатых мышц
Данный тип ткани еще называют скелетной мускулатурой. Волокна этого типа мышц длинные, собранные в пучки. Их клетки могут достигать нескольких сантиметров в длину (вплоть до 10-12). В них содержится много ядер, митохондрий и миофибрилл. Основная структурная единица каждой миофибриллы поперечно-полосатой ткани - саркомер. Он состоит из сократительного белка.
Главная особенность этой мускулатуры заключается в том, что она может контролироваться сознательно, в отличие от гладкой и сердечной.
Волокна данной ткани прикрепляются к костям с помощью сухожилий. Именно поэтому такие мышцы и называются скелетными.
Структура гладкой мышечной ткани
Гладкие мышцы выстилают некоторые внутренние органы, такие как кишечник, матка, мочевой пузырь, а также сосуды. Кроме того, из них формируются сфинктеры и связки.
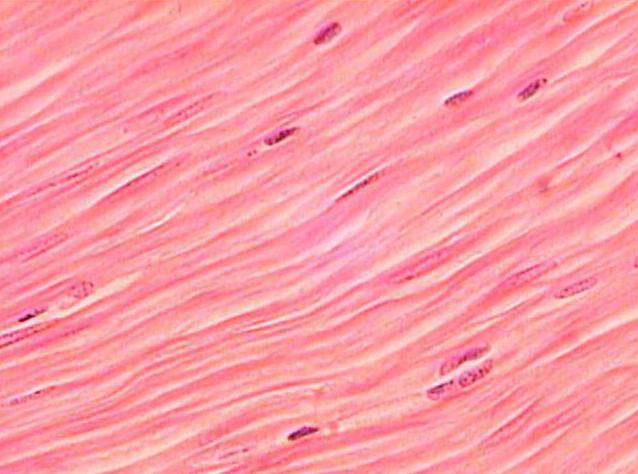
Гладкое не такое длинное, как поперечно-полосатое. Но толщина его больше, чем в случае со скелетными мускулами. Клетки гладкой мышечной ткани обладают веретоноподобной формой, а не нитевидной, как миоциты поперечно-полосатой.
Структуры, которые обесечивают сокращение гладких мышц, называются протофибриллами. В отличие от миофибрилл, они обладают более простой структурой. Но материал, из которого они построены, - все те же сократительные белки актин и миозин.
Митохондрий в миоцитах гладкой мускулатуры также меньше, чем в клетках поперечно-полосатой и сердечной. Кроме того, в них содержится только одно ядро.
Особенности сердечной мышцы
Некоторые исследователи определяют ее как подвид поперечно-полосатой мышечной ткани. Их волокна и вправду во многом похожи. Клетки сердца - кардиомиоциты - также содержат несколько ядер, миофибриллы и большое количество митохондрий. Данная ткань, как и способна сокращаться намного быстрее и сильнее, нежели гладкая мускулатура.
Однако основной особенностью, отличающей сердечную мышцу от поперечно-полосатой, является то, что она не может контролироваться сознательно. Сокращение ее происходит только автоматически, как и в случае с гладкими мышцами.
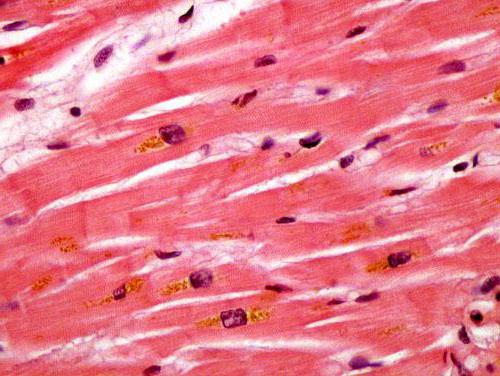
В составе сердечной ткани, кроме типичных клеток, присутствуют также секреторные кардиомиоциты. Они не содержат в себе миофибрилл и не сокращаются. Эти клетки отвесают за выработку гормона атриопептина, который необходим для регуляции артериального давления и контроля объема циркулирующей крови.
Функции поперечно-полосатых мышц
Основная их задача - перемещение тела в пространстве. Также это перемещение частей тела относительно друг друга.
Из других функций поперечно-полосатых мышц можно отметить поддержание позы, депо воды и солей. Кроме того, они выполняют защитную роль, что особенно касается мышц брюшного пресса, предотвращающих механическое повреждение внутренних органов.
К функциям поперечно-полосатой мускулатуры можно также причислить регуляцию температуры, так как при активном сокращении мышц происходит выделение значительного количества тепла. Вот почему при перемерзании мышцы начинают непроизвольно дрожать.
Функции гладкой мышечной ткани
Мускулатура данного вида выполняет эвакуаторную функцию. Она заключается в том, что гладкие мышцы кишечника проталкивают каловые массы к месту их выведения из организма. Также эта роль проявляется при родах, когда гладкие мышцы матки выталкивают плод из органа.
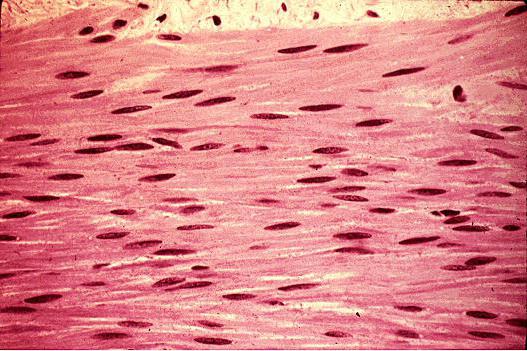
Функции гладкой мышечной ткани этим не ограничиваются. Также немаловажна их сфинктерная роль. Из ткани данного вида формируются специальные круговые мышцы, которые могут смыкаться и размыкаться. Сфинктеры присутствуют в мочевых путях, в кишечнике, между желудком и пищеводом, в желчном пузыре, в зрачке.
Еще одна важная роль, которую играют гладкие мышцы, - формирование связочного аппарата. Он необходим для поддержания правильного положения внутренних органов. При понижении тонуса этих мышц может происходить опущение некоторых органов.
На этом функции гладкой мышечной ткани заканчиваются.
Предназначение сердечной мышцы
Здесь, в принципе, особо говорить не о чем. Основная и единственная функция этой ткани - обеспечение циркуляции крови в организме.
Вывод: различия между тремя видами мышечной ткани
Для раскрытия этого вопроса представляем таблицу:
| Гладкая мускулатура | Поперечно-полосатые мышцы | Сердечная мышечная ткань |
| Сокращается автоматически | Может контролироваться сознательно | Сокращается автоматически |
| Клетки удлинненные, веретеноподобные | Клетки длинные, нитевидные | Удлинненные клетки |
| Волокна не объединяются в пучки | Волокна объединяются в пучки | Волокна объединяются в пучки |
| Одно ядро в клетке | Несколько ядер в клетке | Несколько ядер в клетке |
| Сравнительно небольшое количество митохондрий | Большое количество митохондрий | |
| Отсутствуют миофибриллы | Присутствуют миофибриллы | Есть миофибриллы |
| Клетки способны делиться | Волокна не могут делиться | Клетки не могут делиться |
| Сокращаются медленно, слабо, ритмично | Сокращаются быстро, сильно | Сокращаются быстро, сильно, ритмично |
| Выстилают внутренние органы (кишечник, матка, мочевой пузырь), формируют сфинктеры | Крепятся к скелету | Формируют сердце |
Вот и все основные характеристики поперечно-полосатой, гладкой и сердечной мышечных тканей. Теперь вы ознакомлены с их функциями, строением и главными различиями и сходствами.
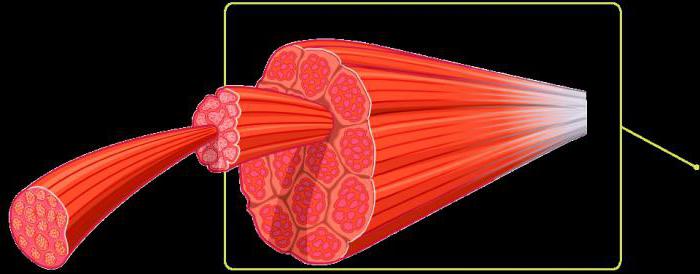
Скелетная мышца состоит из многочисленных пучков мышечных волокон (фасцикул), видимых невооруженным глазом. Один пучок мышечных волокон состоит приблизительно из 10-20 параллельно расположенных мышечных волокон. Каждое мышечное волокно, в свою очередь, содержит 1000-2000 миофибрилл (Silbernagl, Despopoulos, 1983; Klee, 2003). Каждая миофибрилла образована несколькими тысячами последовательно расположенных саркоме (Lindel, 2006), которые являются сократительными элементами мышечного волокна
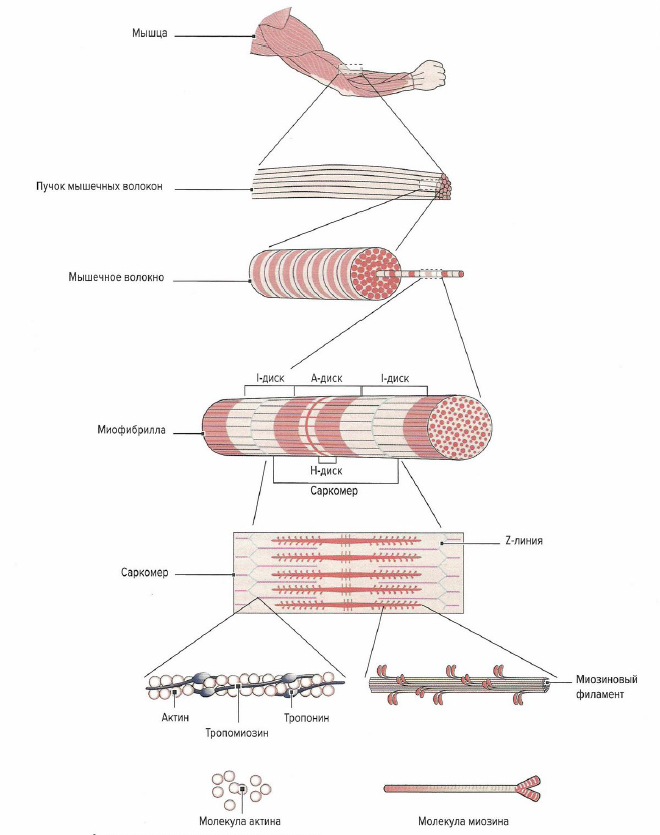
Поперечнополосатое мышечное волокно представляет собой мельчайшую самостоятельную структурную единицу скелетной мышцы. Это одиночная цилиндрическая клетка (см. примечание научного редактора выше), длина которой может достигать от нескольких миллиметров до 30 см (Cabri, 1999). Наиболее длинными мышечными волокнами в организме человека обладает портняжная мышца (m. sar-torius), один из сгибателей бедра. Толщина мышечного волокна зависит от клеточного питания (трофики) и нагрузки на мышцу и варьирует от 10 до 100 мкм. Например, мышцы бедра имеют очень толстые мышечные волокна, тогда как мышцы глазного яблока состоят из тонких волокон (Cabri, 1999). Толщина мышечных волокон в мышце относительно постоянна. Таким образом, брюшко мышцы формируется не из-за утолщения волокон в центре мышцы, а из-за неравной длины мышечных волокон, накладывающихся друг на друга, что образует веретенообразную форму мышцы. Мышечное волокно имеет несколько сотен ядер, объединенных одной клеточной мембраной (сарколеммой) и расположенных в цитоплазме (саркоплазме). В саркоплазме мышечных волокон находятся сократительные миофибриллы, саркоплазматический ретикулум (разновидность эндоплазматического ретикулума), митохондрии (саркосомм), лизосомы, капельки жира и гранулы гликогена. Помимо этого, в саркоплазме растворены молекулы креатинфосфата, аминокислоты, гликолитические ферменты, гликоген и другие вещества. Большую часть саркоплазмы занимают миофибриллы. Они тянутся от одного конца волокна к другому и имеют толщину около 1-2 мкм (Cabri, 1999). При изучении под электронным микроскопом видно, что каждая мио-фибрилла состоит из двух белковых миофиламентов.
- Актиновые миофиламенты — имеют в поперечнике около 5-8 нм (Tillmann, 1998; Klee, 2003). Вокруг актина дополнительно закручены нити троиомиозина, а на равных интервалах (примерно каждые 40 нм) прикрепляются молекулы тропопина (Silbernagl, Despopoulos, 1983). Тропонии (Tn) состоит из трех субъединиц: Tn-С (образует связь с Са2+), Тп-Т (соединяет тропонин с тропо-миозином) и Тп-1 (блокирует образование мосто-видных связей между миозином и актином в состоянии покоя). При соединении субъединицы Тп-С с ионами кальция это тормозящее действие прекращается и актиновые филаменты прикрепляются к Z-линии.
- Миозиновые филаменты — расположены между филаментами актина. Миозиновые филаменты образованы из пучка плотно соединенных молекул миозина (около 150-360) и составляют 1,6 мкм в длину и 10-14 мкм в толщину. Молекулы миозина имеют двойные головки, прикрепляющиеся с помощью шейки к шарнирному участку (тяжелый меромиозин). Шейка молекулы обеспечивает ее подвижность и переходит в длинную хвостовую нить (легкий меромиозин). Благодаря шарнирным участкам миозин может образовывать обратимые связи с актином и вызывать взаимное скольжение нитей актина и миозина относительно друг друга.
Отрезок между Z-линиями называется саркоме-ром. Микроскопически саркомер состоит из чередующихся светлых и темных полосок и линий, обусловленных упорядоченным расположением толстых нитей актина и тонких нитей миозина. Благодаря различному преломлению света актиновыми и миозиновыми филаментами формируется характерная поперечная исчерченность мышечного волокна. Актиновые филаменты в меньшей степени преломляют свет (изотропные) и выглядят как светлые линии, в то время как сильно преломляющие свет миозиновые фила-менты (анизотропные) выглядят темными полосками под микроскопом. Отрезок между двумя соседними саркомерами, содержащий только нити актина, называется 1-диском. В середине 1-диска видна темная линия — Z-линия — граница саркомера. Между двумя 1-дисками расположен A-диск, состоящий как из миозина, так и из актина. В середине A-диска в состоянии расслабления виден еще один светлый промежуток — Н-диск, — содержащий только миозин. При сокращении нити актина сдвигаются к центру миозинового филамента и Н-диск исчезает. Длина саркомера в состоянии покоя, по данным литературы, варьирует и составляет от 2,0 (Trombitas et al, 1998) до 2,2 мкм (Klee, 2003).
Помимо актина и миозина в мышечном волокне содержатся и другие белки (рис. 1.2). В отличие от вышеперечисленных белков они не относятся к сократительным. Их функцией является поддержание структурной организации внутренних компонентов мышечного волокна, благодаря чему достигается стабилизация волокна во всех плоскостях и его внутренние компоненты могут выдерживать внешние и внутренние нагрузки как в продольном, так и в поперечном направлении. Эти белки образуют следующие третичные филаменты (Maruyama etal., 1984; Street, 1983).
- Небулиновые филаменты — состоят из белка небулина и расположены параллельно тонким актиновым нитям. Небулиновые филаменты прикрепляются к Z-линиям и стабилизируют актиновые филаменты и контролируют расположение молекул актина (Cabri, 1999).
- Хитиновые филаменты (коннектиновые филаменты) — состоят из одной молекулы белка титина. Данные филаменты длиной 1 нм в натянутой мышце занимают половину саркомера, соединяя М-линию и Z-линию (Maruyama etal., 1984). Титиновые филаменты расположены параллельно нитям миозина и прикрепляются к их свободным концам. Миозиновый комплекс, соединяясь с 3-6 хитиновыми филаментами, образует функциональную единицу (Klee, 2003). Ее функцией является удерживание миозиновых филаментов при сокращении в центре саркомера. Между Z-линией и концами нитей миозина они образуют высокоэластичную PEVK-область [образуемую пролином (Pro или Р) глутаминовой кислотой (Glu или Е), валином (Val или V) и лизином (Lys или К); Linke et al. 1996]. Благодаря наличию этого эластичного участка саркомер может восстанавливать свою изначальную длину (длину в покое) после растяжения за счет притягивания миозиновых филаментов к Z-линиям. Это позволяет восстановить оптимальное взаиморасположение актиновых и миозиновых нитей. Таким образом, титиновые филаменты ответственны за остаточное напряжение мышц при их расслаблении (тонус мышцы) (Wiemann et al, 1998). Промежуточные филаменты — расположены перпендикулярно направлению сократительных филаментов. а-Актин спиралеобразно окружает Z-линию и участвует в стабилизации актиновых филаментов. Белок десмин прикрепляется к сарколемме и соединяет друг с другом соседние миофибриллы, стабилизируя таким образом мышечное волокно в поперечном направлении (Street, 1993; Wang, 1984). Это позволяет сохранить структуру А- и I-дисков и поперечную испорченность мышечных волокон.
- Поперечно расположенные М-белки, миомезин и М-креатинкиназы (М-КК) образуют в центре Н-диска так называемую М-линию. Миомезин при этом играет функцию якоря для хитина, М-КК обеспечивает образование АТФ, а М-белок соединяет друг с другом и удерживает миозиновые филаменты.
- С-белки образуют С-линии. Продольно расположенные филаменты из С-белка стабилизируют миозиновые нити в саркомере.
- Короткие филаментные и глобулярные белки — расположены в сарколемме (а- и p-интегршг), в саркоплазматической мембране (дистрофии, талин, винкулин) и вне сарколеммы (ламинин и фибронсктин), контактируя с оболочкой сократительных волокон.
Запомните:
Саркомер состоит из тонких актиновых и толстых миозиновых филаментов, а также несократительных «третичных филаментов». Несколько тысяч последовательно расположенных саркомеров образуют миофибриллу. Большое количество рядом расположенных миофибрилл, в свою очередь, формирует мышечное волокно. Параллельно расположенные мышечные волокна образуют пучок мышечных волокон (фасцикулу), из которых состоит анатомически определяемая мышца.
Миофибриллы окружены эндоплазматическим ретикулумом, который в мышцах называется саркоплазматическим ретикулумом. Он представляет собой систему продольно расположенных плоских разветвленных трубочек, соединенных между собой (L-система, продольная система) и отграниченных от сарколеммы и межклеточного пространства.
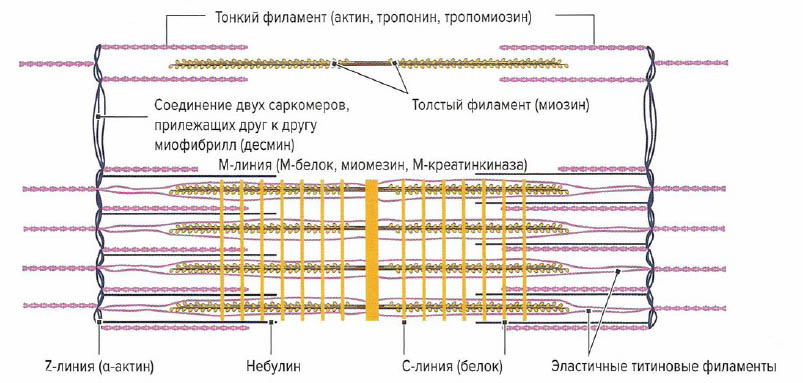
Около Z-лииий на протяжении всей миофибриллы саркоплазматический ретикулум образует мешковидные расширения (терминальные цистерны). Эта органелла играет роль резервуара ионов кальция и ответственна за их накопление и обратный захват. К саркоплазматическому ретикулуму также можно отнести поперечную систему (Т-система, Т-трубочки). Под поперечными трубочками понимают систему вертикально расположенных выпячиваний сарколеммы, соединяющих ее с экстрацеллюлярным пространством. Данные выпячивания диаметром около 50 нм располагаются по бокам от Z-линий (Forssmann, 1985). Они выполняют роль ионных каналов, которые открываются и закрываются в зависимости от потенциала действия. Кроме этого, они обусловливают быстрое проведение потенциала действия от поверхности клеточной мембраны к центру клетки и одновременное сокращение всех миофибрилл внутри одного мышечного волокна. В определенных местах поперечные трубочки располагаются между двумя L-цистернами продольной системы, образуя так называемую триаду. За счет накопления мембранного потенциала в щелевидных контактах (T-L-соединение) происходит распространение возбуждения по L-системе всей мышечной клетки, что вызывает высвобождение ионов кальция.
Другими важными компонентами саркоплазмы являются митохондрии, в мышечных волокнах называемые саркосомами. Количество саркосом варьирует в зависимости от типа мышечных волокон. Саркосомы находятся между миофибриллами или непосредственно под саркосомой. В центре волокна саркосомы расположены либо циркулярно вокруг Z-линии, либо между миофибриллами. Между миофибшеллами в виде «ожерелий» находятся маленькие гранулы гликогена. Их также в больших количествах обнаруживают около клеточного ядра. Часто около митохондрий в саркоплазме видны липиды в виде маленьких капель. Количество липидов в мышечном волокне зависит от его типа.
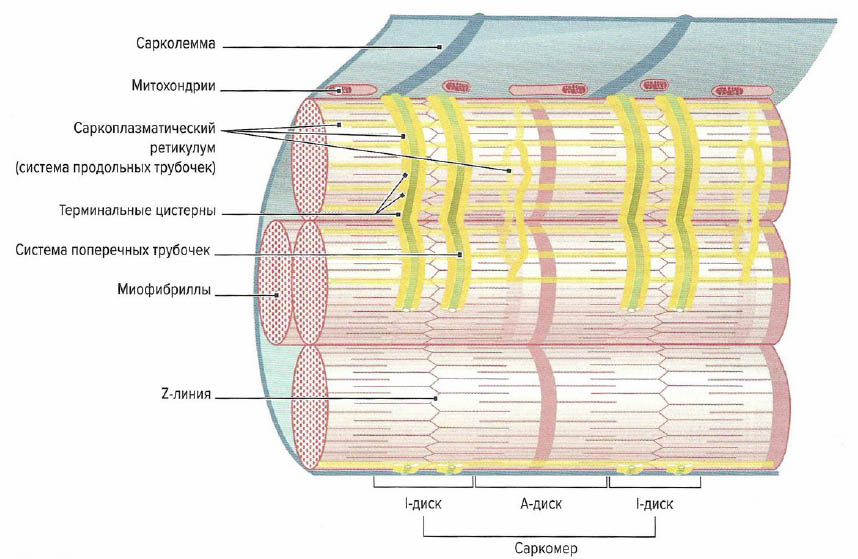
Сократительные элементы мышечных волокон и все компоненты саркоплазмы окружены клеточной мембраной (сарколеммой). Сарколемма представляет собой эластичную способную к возбуждению мембрану, функцией которой является открытие и закрытие ионных каналов в ответ на возбуждающие стимулы и их дальнейшая передача по Т-системе. Сарколемма, в свою очередь, окруокена базальной мембраной, которая отделяет мышечные волокна от соединительной ткани мышцы.
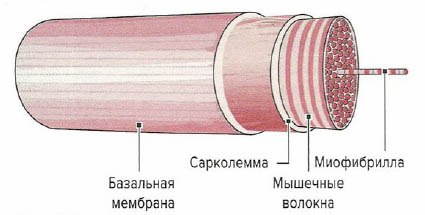
Помимо сократительных элементов скелетные мышцы состоят из соединительной ткани» окружающей мышцы и содержащей кровеносные сосуды и нервы.
Последние материалы сайта
Наследственные

«Прекрасный фонтан» на центральной площади Нюрнберга
Фотографии Антона Дергачева (кроме первой), комментарий Владимира Дергачева Идеальная супружеская пара на лебедях, которая вот-вот поцелуются. Фонтан «Супружеская или Брачная карусель» создан по мотивам немецкого средневекового поэта и композитора Ганс
Симптомы

Карта-схема комплексного обследования
Логопедическое обследование детей с церебральным параличом Логопедическое обследование детей, страдающих церебральным параличом, строится на общем системном подходе, который опирается на представление о речи как о сложной функциональной системе, структ
Методы диагностики

Пять компотов из вишни на зиму
Александр Гущин За вкус не ручаюсь, а горячо будет:) Содержание Яркая, красивая ягода вишня содержит в себе огромное количество полезных микроэлементов и витаминов. Ее универсальность в кулинарии позволяет приготовить массу хороших, вкусных, уникаль
Болевые синдромы

Готовим изумительную пшенную кашу на молоке в мультиварке редмонд Как сварить пшенную кашу скороварке
Ни одна кастрюля в мире не сварит пшенную кашу на молоке так вкусно, как это сделает мультиварка. И ни одна плита не справится с ее приготовлением так быстро, как мультиварка-скороварка. Простой и здоровый завтрак – сладкая пшенная каша на молоке, сдобрен
Синдромы

Рыцарь монет таро значение в отношениях
Название: Рыцарь пентаклей, Рыцарь денариев, Рыцарь монет, Воин монет, Рыцарь бубен, Путешественник, Завоеватель Дел, Всадник, Кавалер, Полезный человек, Повелитель Земель Диких и Плодородных. Значение по Папюсу: Выгода, Прибыль, Польза, Интерес. Важное
Методы лечения

Гороскоп на август телец девушка
Томный август у многих Тельцов станет благоприятным временем для развития личных отношений. В этот период у представителей данного знака Зодиака возникает возможность использовать на полную катушку свое природное обаяние для успеха в сердечных делах. Чтоб